En blomst er en iøynefallende, ofte vakker, viktig del av blomstrende planter. Blomster kan være store og små, fargerike og grønne, luktende og luktfrie, enkeltstående eller samlet fra mange små blomster til en felles blomsterstand.
En blomst er et modifisert forkortet skudd som tjener til frøreproduksjon. Blomsten ender vanligvis på hoved- eller sideskuddet. Som ethvert skudd utvikler en blomst seg fra en knopp.
blomsterstruktur
Blomst - reproduksjonsorganet til angiospermer, som består av en forkortet stilk (blomstaksen), hvorpå blomsterdekselet (perianth), støvbærere og pistiller, bestående av en eller flere fruktblader, er plassert.
Blomstens akse kalles beholder. Beholderen, som vokser, tar en annen form - flat, konkav, konveks, halvkuleformet, kjegleformet, langstrakt, søyleformet. Beholderen på bunnen går inn i pedicelet, og forbinder blomsten med stilken eller stilken.
Blomster som ikke har pedicel kalles fastsittende. På pedicelet til mange planter er det to eller ett lite blad - bracts.
blomsterdekke - perianth- kan deles i en kopp og en visp.
Kopp danner den ytre sirkelen av perianth, bladene er vanligvis relativt små i størrelse, Grønn farge. Skille mellom separat og leddbladet beger. Vanligvis utfører den funksjonen med å beskytte de indre delene av blomsten til knoppen åpner seg. I noen tilfeller faller begeret av når blomsten blomstrer, oftest forblir den under blomstringen.
Delene av blomsten som ligger rundt støvbærerne og pistillen kalles perianth.
De indre bladene er kronbladene som utgjør kronen. De ytre bladene - begerbladene - danner et beger. Perianth, som består av en beger og en krone, kalles dobbel. Perianth, som ikke er delt inn i krone og beger, og alle bladene på blomsten er mer eller mindre like - enkelt.

Corolla- den indre delen av perianth, skiller seg fra begeret i lys farge og større størrelse. Fargen på kronbladene skyldes tilstedeværelsen av kromoplaster. Skille separat - og felles-kronblad corollas. Den første består av individuelle kronblader. I interpetal corollas skilles et rør og en lem vinkelrett på den, som har et visst antall tenner eller skovler av corolla.
Blomster er symmetriske og asymmetriske. Det er blomster som ikke har perianth, de kalles nakne.
Symmetrisk (aktinomorf)- hvis mange symmetriakser kan trekkes gjennom vispen.
Asymmetrisk (zygomorf)- hvis bare én symmetriakse kan tegnes.
Doble blomster har et unormalt økt antall kronblader. I de fleste tilfeller skyldes de splitting av kronbladene.
Stamen- en del av en blomst, som er en slags spesialisert struktur som danner mikrosporer og pollen. Den består av en filament som den er festet til beholderen gjennom, og en støvfang som inneholder pollen. Antall støvbærere i en blomst er et systematisk trekk. Støvbærere kjennetegnes av metoden for feste til beholderen, av formen, størrelsen, strukturen til stamenfilamentene, bindemiddel og støvbærer. Samlingen av støvbærere i en blomst kalles androecium.

filament- den sterile delen av støvbæreren, med en støvbærer på toppen. Filamentet kan være rett, buet, vridd, viklet, ødelagt. I form - hårete, kjegleformet, sylindrisk, flatt, kølleformet. Av overflatens natur - naken, pubescent, hårete, med kjertler. Hos noen planter er den kort eller utvikler seg ikke i det hele tatt.
Anther plassert på toppen av staminatfilamentet og festet til det med et leddbånd. Den består av to halvdeler forbundet med en lenke. Hver halvdel av støvbæreren har to hulrom (pollensekker, kamre eller reir) der pollen utvikler seg.

Som regel er støvfjæren fircellet, men noen ganger blir skilleveggen mellom reirene i hver halvdel ødelagt, og støvfjæren blir tocellet. Hos noen planter er støvbæreren til og med encellet. Det er svært sjelden å se treenighet. I henhold til typen feste til glødetråden er støvknappene faste, bevegelige og svingende.
Støvbrytere inneholder pollen eller pollenkorn.
Strukturen til pollenkornet
Støvkornene som dannes i støvbærerne til støvbærerne er små korn, de kalles pollenkorn. De største når 0,5 mm i diameter, men vanligvis er de mye mindre. Under et mikroskop kan du se at støvpartiklene til forskjellige planter ikke er like. De er forskjellige i størrelse og form.

Overflaten på støvkornet er dekket med forskjellige fremspring, tuberkler. Når du kommer på stemplet, holdes pollenkorn ved hjelp av utvekster og en klebrig væske som frigjøres på stigmaet.
Reirene til den unge støvbæreren inneholder spesielle diploide celler. Som et resultat av meiotisk deling dannes det fire haploide sporer fra hver celle, som kalles mikrosporer for deres svært lille størrelse. Her, i hulrommet i pollenposen, blir mikrosporer til pollenkorn.

Dette skjer som følger: mikrosporekjernen er delt mitotisk inn i to kjerner - vegetativ og generativ. Rundt kjernene er områder av cytoplasmaet konsentrert og to celler dannes - vegetative og generative. På overflaten av den cytoplasmatiske membranen til mikrosporen dannes et veldig sterkt skall fra innholdet i pollenposen, uløselig i syrer og alkalier. Dermed består hvert pollenkorn av vegetative og generative celler og er dekket med to skjell. Mange pollenkorn utgjør pollenet til en plante. Pollen modnes i støvknappene når blomsten åpner seg.
pollen spiring
Begynnelsen av pollenspiring er assosiert med mitotisk deling, som et resultat av at det dannes en liten reproduktiv celle (sædceller utvikler seg fra den) og en stor vegetativ celle (pollenrør utvikler seg fra den).
Etter at pollen på en eller annen måte kommer på stigmaet, begynner spiringen. Den klissete og ujevne overflaten av stigmaet hjelper til med å holde på pollen. I tillegg frigjør stigmaet et spesielt stoff (enzym) som virker på pollenet og stimulerer dets spiring.
Pollenet svulmer, og den begrensende påvirkningen av exinet (det ytre laget av pollenkornkappen) fører til at innholdet i pollencellen sprekker en av porene, gjennom hvilken intina (det indre, poreløse skallet til pollenkornet) buler utover som et smalt pollenrør. Innholdet i pollencellen går inn i pollenrøret.
Under epidermis av stigmaet er løst vev som pollenrøret trenger inn i. Den fortsetter å vokse, og passerer enten gjennom en spesiell ledende kanal mellom de slimete cellene, eller kronglete langs de intercellulære rommene i det ledende vevet i kolonnen. Samtidig går vanligvis et betydelig antall pollenrør frem samtidig i kolonnen, og "suksessen" til ett eller annet rør avhenger av den individuelle veksthastigheten.
To sædceller og en vegetativ kjerne passerer inn i pollenrøret. Hvis dannelsen av spermatozoer i pollen ennå ikke har skjedd, går den generative cellen inn i pollenrøret, og her dannes sædceller ved deling. Den vegetative kjernen er ofte plassert foran, i den voksende enden av røret, og sædceller er suksessivt plassert bak den. I pollenrøret er cytoplasmaet i konstant bevegelse.

Pollen er rik på næringsstoffer. Disse stoffene, spesielt karbohydrater (sukker, stivelse, pentosaner) konsumeres intensivt under pollenspiring. I tillegg til karbohydrater kjemisk oppbygning pollen inkluderer proteiner, fett, aske og en omfattende gruppe enzymer. Pollen inneholder et høyt innhold av fosfor. Stoffer er i pollen i mobil tilstand. Pollen overføres enkelt lave temperaturer opp til - 20º og enda lavere, i lang tid. Høye temperaturer reduserer raskt spiringen.
Støt
Pistillen er den delen av blomsten som danner frukten. Det oppstår fra fruktbladene (en bladlignende struktur som bærer eggene) etter sammensmeltingen av kantene på sistnevnte. Det kan være enkelt hvis det er sammensatt av én fruktblad, og komplekst hvis det er sammensatt av flere enkle pistiller smeltet sammen av sideveggene. Hos noen planter er pistillene underutviklet og representert bare av rudimenter. Pistillen er delt inn i eggstokk, stil og stigma.


Eggstokk- den nedre delen av pistillen, der frøkimene er plassert.
Etter å ha kommet inn i eggstokken, vokser pollenrøret videre og går i de fleste tilfeller inn i eggstokken gjennom polleninntaket (mikropyl). Ved å trenge inn i embryoposen sprekker enden av pollenrøret, og innholdet renner ut på en av synergidene, som blir mørkere og raskt kollapser. Den vegetative kjernen blir vanligvis ødelagt før pollenrøret trenger inn i embryoposen.
Blomster rett og galt
Blomsterbladene (enkle og doble) kan ordnes slik at flere symmetriplan kan trekkes gjennom den. Slike blomster kalles riktige. Blomster som ett symmetriplan kan trekkes gjennom kalles uregelmessige.
Blomster biseksuelle og toboe
De fleste planter har blomster som har både støvbærere og pistiller. Dette er bifile blomster. Men i noen planter har noen blomster bare pistiller - pistillatblomster, mens andre bare har stamens - staminate blomster. Slike blomster kalles dioecious.
Planter eneboende og toboende
Planter som utvikler både pistillate og staminate blomster kalles monoecious. Toebolige planter - staminerer blomster på en plante, og pistillat - på en annen.
Det er arter der bifile og enkjønnede blomster kan finnes på samme plante. Dette er de såkalte polygame (polygame) plantene.
blomsterstander
Blomster dannes på skudd. Svært sjelden befinner de seg alene. Oftere samles blomster i iøynefallende grupper kalt blomsterstander. Begynnelsen på studiet av blomsterstander ble lagt av Linné. Men for ham var blomsterstanden ikke en type forgrening, men en måte å blomstre på.
I blomsterstander skilles hoved- og sideaksene (fastsittende eller på pedikler), da kalles slike blomsterstander enkle. Hvis blomstene er på sideaksene, er dette komplekse blomsterstander.
| Blomsterstand type | Blomsterstandsskjema | Egendommer | Eksempel |
| Enkle blomsterstander | |||
| Børste | Separate sideblomster sitter på en langstrakt hovedakse og har samtidig sine egne pedikler, omtrent like lange | Fuglekirsebær, liljekonvall, kål | |
| Øre | Hovedaksen er mer eller mindre langstrakt, men blomstene er uten stilker, d.v.s. stillesittende. | Plantain, orkide | |
| kolbe | Den skiller seg fra øret i en kjøttfull fortykket akse. | Mais, calla | |
| Kurv | Blomstene er alltid fastsittende og sitter på en sterkt fortykket og utvidet ende av en forkortet akse, som har et konkavt, flatt eller konveks utseende. I dette tilfellet har blomsterstanden utenfor en såkalt omslag, bestående av en eller mange påfølgende rader med bracts, frie eller sammenvoksede. | Kamille, løvetann, aster, solsikke, kornblomst | |
| Hode | Hovedaksen er sterkt forkortet, sideblomstene er fastsittende eller nesten fastsittende, tett plassert i forhold til hverandre. | Kløver, scabiosa | |
| Paraply | Hovedakselen er forkortet; laterale blomster kommer ut, som det var, fra ett sted, sitter på ben av forskjellige lengder, plassert i samme plan eller kuppelformet. | Primula, løk, kirsebær | |
| Skjold | Den skiller seg fra børsten ved at de nederste blomstene har lange stilker, slik at blomstene som et resultat er plassert nesten i samme plan. | Pære, spirea | |
| Komplekse blomsterstander | |||
| Kompleks børste eller panikk |  | Sideforgreningsøkser avviker fra hovedaksen, på hvilken blomster eller enkle blomsterstander er plassert. | Syrin, havre |
| kompleks paraply | Enkle blomsterstander avviker fra den forkortede hovedaksen. | Gulrot, persille | |
| Kompleks øre | Individuelle spikelets er plassert på hovedaksen. | Rug, hvete, bygg, hvetegress | |
Den biologiske betydningen av blomsterstander
Den biologiske betydningen av blomsterstander er at de små, ofte usynlige blomstene, klynget sammen, blir iøynefallende, produserer mest pollen og tiltrekker seg bedre insekter som bærer pollen fra blomst til blomst.
Pollinering
For at befruktning skal skje, må pollenet lande på stemplets stigma.
Prosessen med å overføre pollen fra støvbærerne til stigmaet kalles pollinering. Det er to hovedtyper av pollinering: selvbestøvning og krysspollinering.
selvbestøvning
Under selvbestøvning faller pollen fra en støvbærer på stigmaet til pistillen til samme blomst. Dette er hvordan hvete, ris, havre, bygg, erter, bønner og bomull pollineres. Selvbestøvning i planter skjer oftest i en blomst som ennå ikke har åpnet seg, det vil si i en knopp, når blomsten åpner seg, er den allerede fullført.
Under selvbestøvning smelter kjønnsceller dannet på samme plante sammen og har derfor de samme arvelige egenskapene. Dette er grunnen til at avkommet som er et resultat av prosessen med selvbestøvning er veldig lik foreldreplanten.

krysspollinering
Med kryssbestøvning skjer rekombinasjonen av arvelige egenskaper fra mors og mors organismer, og det resulterende avkommet kan få nye egenskaper som foreldrene ikke hadde. Slike avkom er mer levedyktige. I naturen er krysspollinering mye mer vanlig enn selvbestøvning.

Krysspollinering utføres ved hjelp av ulike eksterne faktorer.
Anemofili(vindpollinering). Hos anemofile planter er blomstene små, ofte samlet i blomsterstander, det dannes mye pollen, det er tørt, lite, og når støvbæreren åpner seg, kastes det ut med kraft. Lyspollen til disse plantene kan bæres av vinden over avstander på opptil flere hundre kilometer.

Støvbærerne er plassert på lange tynne filamenter. Stempelen er brede eller lange, finnede og stikker ut fra blomstene. Anemophilia er karakteristisk for nesten alle gress, sedges.
Entomofi(bærer pollen av insekter). Tilpasningen av planter til entomofili er lukten, fargen og størrelsen på blomster, klissete pollen med utvekster. De fleste blomstene er tvekjønnede, men modning av pollen og pistiller skjer ikke samtidig, eller høyden på stigmaene er større eller mindre enn høyden på støvbærerne, noe som tjener som beskyttelse mot selvbestøvning.
I blomstene til insektpollinerte planter er det områder som skiller ut en søt duftende løsning. Disse områdene kalles nektarier. Nectaries kan være plassert på forskjellige steder av blomsten og har ulike former. Insekter, som har fløyet opp til blomsten, trekkes til nektarier og støvbærere, og under måltidet blir de skitne med pollen. Når et insekt flytter til en annen blomst, fester pollenkornene som den bærer seg til stigmaene.
Ved pollinering av insekter går mindre pollen til spille, og derfor sparer planten stoffer ved å produsere mindre pollen. Pollenkorn trenger ikke holde seg lenge i luften og kan derfor være tunge.
Insekter kan bestøve sparsomt beliggende blomster og blomster på rolige steder – i skogkrattet eller tykt gress.
Vanligvis blir hver planteart bestøvet av flere arter av insekter, og hver pollinerende insektart tjener flere plantearter. Men det er noen typer planter hvis blomster pollineres av insekter av bare én art. I slike tilfeller er den gjensidige samsvaret mellom livsmåtene og strukturen til blomster og insekter så fullstendig at det virker mirakuløst.
Ornitofili(bestøvning av fugler). Det er typisk for noen tropiske planter med fargerike blomster, rikelig med sekresjoner av nektar og en sterk elastisk struktur.
hydrofili(pollinering med vann). observert i vannplanter. Pollen og stigmaet til disse plantene har oftest en filamentøs form.
bestialitet(bestøvning av dyr). Disse plantene er preget av store blomsterstørrelser, rikelig sekresjon av nektarholdig slim, masseproduksjon av pollen og blomstring om natten når de pollineres av flaggermus.
Befruktning
Pollenkornet faller på stemplets stempel og festes til det på grunn av skallets strukturelle egenskaper, samt de klebrige sukkerholdige sekretene fra stempelet, som pollenet fester seg til. Pollenkornet sveller og spirer til et langt, veldig tynt pollenrør. Pollenrøret dannes som et resultat av deling av en vegetativ celle. Først vokser dette røret mellom cellene i stigmaet, deretter stilen, og til slutt vokser inn i hulrommet i eggstokken.
Den generative cellen til pollenkornet beveger seg inn i pollenrøret, deler seg og danner to mannlige kjønnsceller (spermier). Når pollenrøret kommer inn i embryoposen gjennom pollenpassasjen, smelter en av sædcellene sammen med egget. Befruktning skjer og en zygote dannes.
Den andre sædcellen smelter sammen med kjernen til den store sentrale cellen i embryoposen. Således, i blomstrende planter, oppstår to fusjoner under befruktning: den første sædcellen smelter sammen med egget, den andre med den store sentrale cellen. Denne prosessen ble oppdaget i 1898 av den russiske botanikeren, akademikeren S.G. Navashin og kalte den dobbel befruktning. Dobbel befruktning er typisk bare for blomstrende planter.
Zygoten dannet ved fusjon av kjønnsceller deler seg i to celler. Hver av de resulterende cellene deler seg igjen, osv. Som et resultat av flere celledelinger utvikler det seg et flercellet embryo til en ny plante.
Den sentrale cellen deler seg også, og danner endospermceller, der næringsreserver samler seg. De er nødvendige for ernæring og utvikling av embryoet. Frøkappen utvikler seg fra integumentet til eggløsningen. Etter befruktning utvikler det seg et frø fra eggløsningen, bestående av et skall, et embryo og en tilførsel av næringsstoffer.
Etter befruktning flyter de til eggstokken næringsstoffer, og det blir gradvis til en moden frukt. Perikarpen, som beskytter frøene mot uønskede effekter, utvikler seg fra eggstokkens vegger. Hos noen planter tar også andre deler av blomsten del i dannelsen av frukten.

Sporedannelse
Samtidig med dannelsen av pollen i støvbærerne, skjer dannelsen av en stor diploid celle i eggløsningen. Denne cellen deler seg meiotisk og gir opphav til fire haploide sporer, som kalles makrosporer fordi de er større enn mikrosporer.

Av de fire dannede makrosporene dør tre av, og den fjerde begynner å vokse og blir gradvis til en embryosekk.
Formasjon av embryosekk
Som et resultat av tredelt mitotisk deling av kjernen i hulrommet i embryoposen, dannes åtte kjerner som er kledd med cytoplasma. Det dannes celler uten membraner, som er ordnet i en bestemt rekkefølge. Ved den ene polen av embryoposen dannes et eggapparat, bestående av et egg og to hjelpeceller. På motsatt pol er det tre celler (antipoder). En kjerne migrerer fra hver pol til midten av embryoposen (polare kjerner). Noen ganger smelter polarkjernene sammen og danner den diploide sentrale kjernen i embryoposen. Embryosekken der kjernefysisk differensiering har skjedd, anses som moden og kan akseptere sædceller.

Når pollenet og embryoposen har modnet seg, åpner blomsten seg.
Strukturen til eggløsningen
Eggløsningene utvikler seg videre indre sider veggene i eggstokken og, som alle deler av planten, består av celler. Antall eggløsninger i eggstokkene til forskjellige planter er forskjellig. I hvete, bygg, rug, kirsebær inneholder eggstokken bare en eggløsning, i bomull - flere dusin, og i valmue når antallet flere tusen.
Hver eggløsning er dekket med et dekke. På toppen av eggløsningen er det en smal kanal - polleninngangen. Det fører til vevet som opptar den sentrale delen av eggløsningen. I dette vevet, som et resultat av celledeling, dannes en embryosekk. På motsatt side av polleninngangen er det et egg i den, og den sentrale delen er okkupert av en stor sentral celle.
Utvikling av angiospermer (blomstrende) planter

Dannelse av frø og frukt
Under dannelsen av et frø og et foster smelter en av sædcellene sammen med egget og danner en diploid zygote. Deretter deler zygoten seg mange ganger, og som et resultat utvikler et flercellet embryo av planten. Den sentrale cellen, som har smeltet sammen med den andre sædcellen, deler seg også mange ganger, men det andre embryoet vises ikke. Et spesielt vev dannes - endospermen. Endospermcellene akkumulerer reserver av næringsstoffer som er nødvendige for utviklingen av embryoet. Integumentene til eggløsningen vokser og blir til en frøkappe.

Som et resultat av dobbel befruktning dannes det således et frø, som består av et embryo, lagringsvev (endosperm) og frøskal. Fra eggstokkveggen dannes fruktveggen, kalt perikarp.
seksuell reproduksjon
Seksuell reproduksjon av angiospermer er assosiert med en blomst. Dens viktigste deler er støvbærere og pistiller. De gjennomgår komplekse prosesser knyttet til seksuell reproduksjon.
Hos blomstrende planter er mannlige kjønnsceller (spermier) svært små, mens kvinnelige kjønnsceller (ovuler) er mye større.
I støvbærerens støvknapper oppstår celledeling, noe som resulterer i dannelse av pollenkorn. Hvert pollenkorn av angiospermer består av vegetative og generative celler. Pollenkornet er dekket med to skjell. Det ytre skallet er som regel ujevnt, med pigger, vorter, utvekster i form av et nett. Dette hjelper pollenkornene til å holde seg til stemplets stigma. Plantens pollen, som modnes i støvbærerne, når blomsten åpner seg, består av mange pollenkorn.
blomsterformel
Formler brukes til å betinget uttrykke strukturen til blomster. For å lage en blomsterformel brukes følgende notasjon:
En enkel perianth, bestående av begerblad eller kronblad alene, dens deler kalles bladblader. |
|
| H | Beger sammensatt av begerblader |
| L | Corolla, sammensatt av kronblader |
| T | Stamen |
| P | Støt |
| 1,2,3... | Antall blomsterelementer er angitt med tall |
| , | De samme delene av en blomst, forskjellige i form |
| () | sammenvoksede deler av en blomst |
| + | Arrangement av elementer i to sirkler |
| _ | Øvre eller nedre eggstokk - en strek over eller under tallet som viser antall pistiller |
| feil blomst | |
| * | høyre blomst |
| ♂ | Enkjønnet utholdenhetsblomst |
| ♀ | enkjønnet pistillatblomst |
| Bifil | |
| ∞ | Antall blomsterdeler større enn 12 |
Eksempel på kirsebærblomstformel:
*H 5 L 5 T ∞ P 1
blomsterdiagram
Strukturen til en blomst kan uttrykkes ikke bare med en formel, men også med et diagram - en skjematisk representasjon av en blomst på et plan vinkelrett på blomstens akse.

Lag et diagram av tverrsnitt av uåpnede blomsterknopper. Diagrammet gir en mer fullstendig ide om strukturen til blomsten enn formelen, siden den også viser den relative plasseringen av delene, som ikke kan vises i formelen.
5. klasse
Utstyr: blomst layout; gul og grønn plastelina, aluminium og kobbertråd, papiremner for begerblad og kronblad.
Når du studerer dette materialet i 5. klasse, er målet å gjøre elevene kjent med strukturen til en blomst og sikre assimilering av begrepene "perianth", "hoveddeler av en blomst". Den foreslåtte praktiske delen utvikler evnen til å observere og sammenligne og skaper forutsetninger for opplæring av nøyaktighet.
UNDER KLASSENE
I. Organisatorisk øyeblikk
II. Sjekker lekser
Hva er et organ? Nevn organene til en blomstrende plante og beskriv deres funksjoner.
(Kontrollere riktigheten av å fylle ut tabellen over organer og funksjoner til en blomstrende plante på s. 160, § 40.)
III. Oppdatering av grunnleggende kunnskap
For hva blomsterplanter fått navnet deres? Hva er et annet navn på disse plantene? Hvorfor?
IV. Lære nytt stoff
Læreren navngir delene av blomsten, forteller om deres struktur og funksjoner. Elevene i gruppen, etter å ha lyttet til forklaringen, konstruerer den tilsvarende delen av blomsten av plastelina og skriver ned navnet i en notatbok. Som et resultat setter hver gruppe sammen en blomstermodell.
Skrive i en notatbok

V. Konsolidering
List opp delene av en blomst. Finn dem på modellene dine.
Hva er hoveddelene av en blomst? Hvorfor?
Hva er støvbærere og pistiller laget av?
Hva er en perianth? Hva er dens funksjoner?
Tegn en blomst i notatboken som ovenfor og ordne tallene for delene av blomsten i riktig rekkefølge.
VI. Hjemmelekser
Studer tilsvarende avsnitt, notater og tegning i notatboken.
6. klasse
Utstyr: tekster (se vedlegg) - en per gruppe; blomstermodeller laget i 5. klasse; lysbilder - på gjennomsiktig film eller i elektronisk form og et middel for demonstrasjon (overheadprojektor, datamaskin med videoutgang, TV eller multimediaprojektor).
Når du studerer dette materialet i 6. klasse, er målet å utvide og konsolidere kunnskap om blomsten, for å sikre assimilering av kunnskap om klassifisering av blomster. Arbeidet bidrar til utvikling av evnen til å sammenligne, analysere og trekke konklusjoner; bidrar til utdanning av en kommunikativ kultur, en følelse av skjønnhet, utvikling av kognitiv interesse for faget.
UNDER KLASSENE
I. Organisatorisk øyeblikk
Klassen er delt inn i grupper på 5-6 personer.
II. Oppdatering av grunnleggende kunnskap
Vi fortsetter å studere den blomstrende planten. Hvilke planter kalles blomstrende planter? Hva er et annet navn for blomstrende planter? Hvorfor? Nevn organene til en blomstrende plante, men husk først hvilke to grupper de er delt inn i.

Forklar disse navnene.
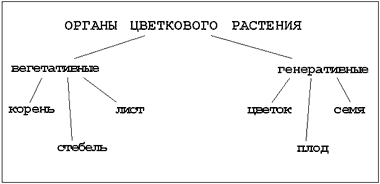
Hvilke organer er vegetative? Hva er en flukt?
Fortsett frasen: organer kalles generative ...
III. Lære nytt stoff
Oppgaven med dagens leksjon er å utvide de eksisterende ideene om blomsten.
Notatbokoppføring:"Blomst".

Blomstene til forskjellige planter skiller seg fra hverandre i størrelse: foran deg er den største blomsten - rafflesia (ca. 1 m i diameter) og den minste - andemat (hele planten passer på miniatyrbildet). Formen og fargen på blomster, antall deler og strukturelle egenskaper er forskjellige. Men det er også likheter.
Når du leser teksten i vedlegget, vil du møte konsepter som du allerede er kjent med. Gjenta dem igjen. studerer nytt materiale tenk på følgende spørsmål.
Hvilke grupper kan blomster deles inn i avhengig av strukturen til perianthen deres?
Hvilke grupper kan blomster deles inn i ved tilstedeværelsen av hoveddelene deres?
(Elevene jobber med tekst.)
V. Kontrollere innledende forståelse
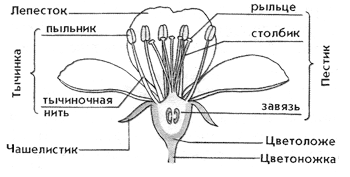
Nevn delene av en blomst, beskriv kort egenskapene til deres struktur og funksjoner.
Hva er en perianth?
Hva er perianter?
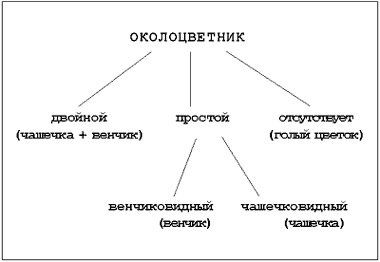
Notatbokoppføring: perianth: dobbel (beger + krone); enkel (kroneformet; begerformet); fraværende (naken blomst).
Hva er hoveddelene av en blomst? Hvorfor?
Hvilke grupper deles blomster inn i etter tilstedeværelsen av hoveddelene deres?

Notatbokoppføring: blomst (i henhold til tilstedeværelsen av hoveddelene): bifil (støvbærer + pistil); tvebolig (staminat); (pistillat); aseksuell (ingen støvbærere eller pistiller)
Hva er ene- og toeboende planter?
V. Konsolidering og anvendelse av ny kunnskap
Og nå skal vi prøve å bruke kunnskapen vi har fått i dagens leksjon i praksis. La oss jobbe med blomstermodeller.
Se på blomstermodellen du laget i 5. klasse og finn ut, basert på tilstedeværelsen av hoveddelene og strukturen til perikarpen, hvilken blomst som er foran deg? ( Bifil med dobbel perianth.)
Hver gruppe studenter mottar et kort med en oppgave, ifølge hvilken det er nødvendig å endre den originale blomsten.
Oppgaver:
1. gruppe - en bifil blomst med en enkel kroneformet perianth;
2. gruppe - bifil blomst med en enkel calyx perianth;
tredje gruppe - en kvinnelig blomst med en dobbel perianth;
Fjerde gruppe - hannblomst med dobbel perianth;
5. gruppe - kvinnelig (eller mannlig) naken blomst;
6. gruppe - aseksuell blomst.
Hver gruppe viser frem sin blomst og forklarer hvilke deler av modellen de har fjernet og hvorfor. De tilsvarende lysbildene vises samtidig.
Spørsmål ( etter demonstrasjon av blomstermodeller av 3. og 4. gruppe): hvilke av gresskarblomstene - hunn eller hann - kalles golde blomster? Hvorfor?
VI. Hjemmelekser
Les det aktuelle avsnittet.
Skriv ned formelen til en vanlig bifil blomst i en notatbok, som har 5 ikke-forente begerblader, 5 ikke-forente kronblader, 5 støvbærere, 1 pistill.
Lag et kryssord om emnet "Blomst" (valgfritt).
Applikasjon.
Tekst til leksjonen
Selv den store tyske poeten Goethe foreslo å betrakte blomsten som et modifisert skudd. Faktisk er det en likhet i strukturen til blomsten og skuddet: pedicel og beholderen er stilkdelen av blomsten, og beger, krone, støvbærere og pistiller er dannet av modifiserte blader.
Således er en blomst et modifisert forkortet skudd som tjener til frø (seksuell) reproduksjon. Som ethvert skudd utvikler en blomst seg fra en knopp. Blomsten ender vanligvis på hoved- eller sideskuddet.
BLOMSTERSTRUKTUR
Beholder - blomstens akse, som, i vekst, kan få en annen form: flat, konkav, konveks osv. Beholderen under går inn i pedicel - en tynn stilk som en blomst sitter på i de fleste planter. På pediselen utvikler mange planter to (i tofrøbladede) eller ett (i enfrøbladede) små blader - dette er stipulerer. En blomst som ikke har en pedicel kalles sittende.
Perianth , som består av kopper Og krone kalt dobbelt. En slik perianth finnes i kirsebærblomster, kål, roser og mange andre planter.

Kopp
danner periantens ytre sirkel. Begeret består vanligvis av små grønne blader - begerblader(H). Hos noen planter, som nellik, vokser begerbladene sammen til et rør med de nedre delene - en slik beger kalles leddblad. Hos andre, som pelargonier, vokser ikke begerbladene sammen, de enkeltbladet kopp.
Når blomsten blomstrer, faller i noen tilfeller begeret av, men oftere forblir det under blomstringen.
Koppen utfører følgende funksjoner:
1) beskytter de indre delene av blomsten til knoppen åpner seg.
2) i de grønne begerbladene finner prosessen med fotosyntese sted.
Corolla
- den indre delen av perianth, består vanligvis av fargesterke store kronblader(L). Hos noen planter (duftende tobakk, svart nattskygge, primula) vokser kronbladene til kronen sammen og danner sympatisk Visp; i andre (kål, eple, kirsebær) består kronen av individuelle kronblader og kalles separat kronblad, eller gratis kronblad.
Hos noen planter, hovedsakelig hos monokotblader (lilje, amaryllis, tulipan), er alle kronbladene mer eller mindre like. En slik perianth kalles enkel(OM). I noen planter, for eksempel i en tulipan, hasselrype eller orkide, er bladene til en enkel perianth store og lyse, som kronblader, - disse er enkel kronblad.

I andre planter, for eksempel i rush, rødbeter, brennesle, er bladene til en enkel perianth små, usynlige, vanligvis grønne, ligner begerblader - dette enkel begerperianth.
Hovedfunksjonene til visp:
1) tiltrekning av pollinerende insekter;
2) beskyttelse av hoveddelene av blomsten.
Det er blomster som ikke har perianth (aske, sedge, selje), kalles de naken.
Blomsterbladene (enkle og doble) kan ordnes slik at flere symmetriakser kan trekkes gjennom dem (eple, kirsebær, kål, etc.). Slike blomster kalles riktig. Blomster som en symmetriakse (erter, salvie) eller ingen (canna) kan trekkes gjennom, kalles feil.
Støt (P) og støvbærere (T) - hoveddelene av en blomst, danner de kjønnsceller - kjønnsceller.
Stamen omfatter filament, som den er festet til beholderen med, og støvfangeren, som inneholder pollen med mannlige kjønnsceller - sperm. Hvis det ikke er noen filament og støvbæreren er plassert direkte på beholderen, kalles det stillesittende.

Støt delt inn i stigma(øverste del laget av spesialstoff, tjener til å fange pollen), kolonne Og eggstokk(nedre utvidet del, der kvinnelige gameter - egg) modnes. Hvis det ikke er stil i pistilen og stigmaet er plassert på eggstokken, kalles det stillesittende. En frukt med frø utvikler seg fra blomstens pistill.

De fleste planter har blomster som har både støvbærere og pistiller. Dette bifile blomster. Men i noen planter (agurk, mais) har noen blomster bare pistiller - dette pistillat, eller hunn, blomster, og andre er bare støvbærere, disse er utholde, eller menns, blomster. Slike blomster kalles tobolig.
aseksuell kalt slike blomster, som mangler alle hoveddelene: både støvbærere og pistiller. De utfører bare funksjonen til å tiltrekke pollinerende insekter til andre blomster der hoveddelene er til stede. Aseksuelle blomster finnes i blomsterstandene til kornblomst, solsikke og andre planter.
Noen blomster har nektarier- kjertler som produserer en søt væske. Oftest er de plassert på beholderen.
Formler brukes til å symbolisere strukturen til en blomst. Følgende notasjon brukes til å formulere formelen:
O - enkel perianth;
H - begerblader;
L - kronblad:
T - støvbærere;
P - støder.
Antall begerblader, kronblader, støvbærere, pistiller er vist i tall, og hvis det er mer enn tolv, så med et tegn.
Hvis noen deler av blomsten har vokst sammen, er de tilsvarende tallene skrevet i parentes.
Den riktige blomsten er avbildet med en stjerne *;
feil-pil;
mannlige (utholdende) blomster av samme kjønn -;
kvinnelig (pistil) - tegn;
bifil - tegn.
bifil kalles en blomst hvor det er både pistiller og støvbærere (androecium og gynoecium). Noen ganger brukes begrepene også på den bifile blomsten. perfekt eller monoecious blomst.
En blomst som bare har støvbærere (androecium) eller bare pistiller (gynoecium) kalles homofil. Enkjønnede blomster med støvbærere utholde, eller menns blomster; følgelig blomster bare med pistiller - pistillat, eller kvinnelige blomster.
Hann- og hunnblomster av samme kjønn kan vokse på samme plante, da kalles planten monoecious, eller bifil, For eksempel: eik, bjørk, spurge, mais. I dette tilfellet kan pollinering forekomme mellom blomster innenfor samme plante.
Hvis hann- og hunnblomster vokser videre forskjellige planter, da har vi å gjøre med tobolig anlegg. En toboplante med utholdende blomster kalles mann , og med kvinner - feminin plante, for eksempel: poppel, selje, hamp, brennesle. For befruktning av dioecious arter er tilstedeværelsen av minst to planter av forskjellige kjønn nødvendig - hann og hunn.
En plante som har både tokjønnede og enkjønnede blomster kalles polygam, for eksempel finnes et slikt nabolag i blomsterstandene til Compositae.
Blomster som mangler sporebærende organer er steril, eller aseksuell blomster, som sivblomster i Compositae-blomsterstander.
Et eksempel på en polygam plante: i gerbera-blomsterstanden på bildet er det hannblomster (med gule støvknapper), hunnblomster (med hvite pistiller) og sterile sivblomster langs kanten.
Blomster av korn, sedge.
Blomster av korn, sedge.
Kornblomster er vanligvis små og lite iøynefallende. De er tilrettelagt for vindpollinering, og mangler derfor perianth, siden de ikke har noen grunn til å tiltrekke seg insekter. Kornblomster er plassert på sideskuddene til spikelet og består av støvbærere Og eggstokk Med stigma grener . Blomsten er beskyttet av toppen og bunnen lemmas . To små fargeløse skjell vokser over blomstrende skjell - den såkalte blomsterfilmer , eller lodiculae . Under blomstringen stikker lange støvbærere utover skjellene og sprer pollen i vinden. Kornblomster kan være bifile eller enkjønnede, noen ganger innenfor samme blomsterstand.
Hjørneblomster er også små og lite iøynefallende, samlet i forskjellige aks og sitter i akslene på bracts, kalt dekke vekter . Selve sirblomsten består av støvbærere Og haviazi Med stigma grener . Blomster er bifile og enkjønnede, med og uten perianth. Hårblomsten kan bestå av et sett med skjell, hårete eller frynsede bust eller silkeaktige hår, og er mer vanlig tilstede i biseksuelle eller kvinnelige blomster.
ANDROECIUM
(gr. "mannens hus"): satt mikrosporofyller, støvbærere som består av en filament med en delt i to halvdeler anther inneholder fire mikrosporangia (pollensekk). Støvbærerne er ordnet i en eller to sirkler. Støvdragere er delt inn i frie og smeltede.
Eksistere forskjellige typer androecium, kjennetegnet ved antall smeltede grupper av støvbærere:
-unifraternal(støvbærere i en gruppe, lupin, kamelia),
-bibroderlig(to grupper støvbærere),
-polybroderlig(flere grupper, magnolia, johannesurt),
-broderlig(ikke-forente støvbærere).
Også støvbærere varierer i lengde: lik, ulik, dobbel styrke(av fire støvbærere er to lange), tre sterke(av seks støvbærere er tre lange), fire sterke(av seks støvbærere er fire lange).
Stamen omfatter støvbærervev, i den øvre enden av dette er anther, og den nedre enden er festet til beholderen. Hovedvevet til filamentet er parenkym. Viktige prosesser finner sted i støvfanget - mikrosporogenese(dannelse av mikrosporer i mikrosporangia) og mikrogametogenese(dannelse fra mikrosporer av den mannlige gametofytten). Den sterile støvbæreren kalles staminoder.
Fig.3 Stenbærer- og støvbærerutvikling

Anther består av homogene celler omgitt av epidermis.
Diagram– Dette er en skjematisk projeksjon av en blomst på et plan der blomsten skjærer seg på tvers, vinkelrett på dens akse. Diagramdesignregel: blomsterstandsakse øverst, dekkblad nederst. Symboler på diagrammet: deler av perianth er indikert med buer, begerblader - med et fremspring i midten av buen, kronblader - uten fremspring. Støvbærerne er indikert i form av et tverrsnitt av støvknapp eller filament. Gynoecium - i form av et tverrsnitt av eggstokken. Hvis individuelle medlemmer vokser sammen, er dette indikert på diagrammet med buer.
Androecium
Androecium er en samling støvbærere der mikrosporogenese, mikrogametofytogenese og dannelse av hannsporer forekommer.
Ved ontogeni kan støvbærere legges i form av tuberkler av en vekstkjegle, som i akropetal(dvs. fra bunnen til toppen), og inn basipetal(topp til bunn) sekvens. I det første tilfellet er de yngste støvbærerne plassert nærmere midten av blomsten, og de gamle er nærmere periferien, og i det andre tilfellet, omvendt. Støvbærerne kan være frie eller vokse sammen på ulike måter og i varierende grad. For eksempel, i den tropiske meliaceae-familien, vokser alle 10 støvbærerne sammen med filamentene til et rør ( unifraternal androecium). I johannesurt vokser støvbærerne sammen i bunter, for Compositae holder støvbærerne seg sammen. Hos mange medlemmer av belgfruktfamilien vokser 9 støvbærere sammen, og en forblir fri (den såkalte bibroderlig androecium).
Hver stamen består av en innsnevret filiform eller sjelden båndlignende eller petaloid del - støvbæreren filament og vanligvis en utvidet del - støvbæreren. Anther har to halvdeler koblet til hverandre forbindelse, som er en fortsettelse av filamentet. Forbindelsen blir noen ganger videreført inn i epikoblingen, synlig som et lite fremspring over støvbæreren.
Dannelsen av tråden begynner senere enn støvbæreren, og dens videre forlengelse utføres på grunn av interkalær vekst. Antall tuberkler som kommer ut er noen ganger mindre enn antall støvbærere, senere deler tuberkler seg, og det kan være ganske mange støvbærere (mimosa). Lengden på filamentene varierer fra plante til plante. Oftere er de mer eller mindre like lange som perianten, men noen ganger er de mye kortere eller mange ganger lengre enn den, som for eksempel i det velkjente tropiske området. medisinsk plante nyre-te, eller kattens værhår, fra kjønnsleppefamilien. På et tverrsnitt gjennom filamentet kan man se at det meste består av parenkymalt vev, og en karbunt passerer i midten.
Hver halvdel av støvbæreren har to (sjelden ett) reir - mikrosporangia. Anther-reir kalles noen ganger pollensekker. Hos den modne støvfangeren forsvinner for det meste skilleveggene mellom reirene. Utenfor er støvbæreren dekket med epidermis. Rett under epidermis er et lag av celler av den såkalte endothecium, med fortykkede cellevegger. Når membranene til endothecium tørker ut, åpnes reirene til støvbæreren. Dypere ligger 1-3 lag med mellomstore tynnveggede celler. Det innerste laget som forer hulrommet til pollenposene kalles tapetum. Det antas at innholdet i cellene tjener som mat for å utvikle morceller. mikrosporer(mikrosporocytter) og fremmer deres differensiering. Anther-reir er vanligvis fylt med mikrospore-morceller, mikrosporer og moden pollen. Mikrosporer er kjent for å oppstå fra mikrosporocytter som et resultat av meiose, og mikrosporocytter i seg selv oppstår fra noen få celler i archesporium (et pedagogisk vev som fungerer i de tidlige stadiene av utviklingen av anther-reir). Den modne støvfangeren åpner seg på ulike måter: langsgående sprekker, hull, ventiler osv. Samtidig renner pollen ut.
Tegn på struktur, form, posisjon, antall støvbærere, samt typen androecium selv har veldig viktig for taksonomi av blomstrende planter og kunnskap om deres fylogeni.
Hos noen arter mister en del av støvbærerne sin opprinnelige funksjon, blir sterile og blir til såkalte staminoder. Noen ganger forvandles støvbærene til nektarer - de sekretoriske delene av blomsten som skiller ut nektar. Kronbladene, deres deler, deler av pistillen og til og med utvekster av beholderen kan også bli til nektarer eller osmoforer. Nektarer har en rekke former, er vanligvis plassert i dypet av blomsten og kjennetegnes ofte av deres skinnende overflate.
Gynoecium.
Samlingen av fruktblader av en blomst, som danner en eller flere pistiller, kalles gynoecium. fruktblader, eller fruktblader, er strukturer som antas å være relatert i opprinnelse til bladet. Funksjonelt og morfologisk tilsvarer imidlertid ikke bærbladene vegetative blader, men blader som bærer megasporangia, dvs. megasporofyll. De fleste morfologer tror at i løpet av evolusjonen fra flate og åpne fruktblader, brettet langs ( duplikativt) fruktblader. Så vokste de sammen i kantene og dannet en pistill med dens viktigste del - eggstokken, som bærer eggene. Dermed ble det dannet en unik struktur, som ikke finnes i noen andre grupper av planter, som ligner et lukket kar der pålitelig beskyttede eggløsninger utvikler seg. Strukturen til pistillen er ideell for pollinering og befruktning. I eggstokkene, som er i eggstokken, utføres prosessene med megasporogenese og megagametofytogenese.
Pistillen, eller rettere sagt eggstokken, fungerer som et våtkammer som beskytter eggstokkene mot å tørke ut. Dette gjorde angiospermer praktisk talt uavhengige av fuktighetsnivåer. miljø og var en av faktorene for deres omfattende utvikling av tørre territorier. I tillegg dekker stampen pålitelig eggene fra å bli spist av insekter og delvis fra temperatursvingninger.
En pistil dannet av en karpell kalles enkel, fra to eller flere sammensmeltede karpeller - kompleks. En enkel pistil er vanligvis unilokulær; kompleks kan deles inn i reir eller det kan også være enkelt-redet. Polynesting oppstår enten som et resultat av sammensmelting av karpeller, eller på grunn av dannelsen av ytterligere skillevegger - utvekster av eggstokkens vegger.
Stigmaet til pistillen er en struktur unik og karakteristisk bare for blomstrende planter, designet for å motta pollen. Det utvikler seg på toppen av stilen eller direkte på eggstokken - et fastsittende stigma; sjeldnere (hos arkaiske arter) - langs de smeltede kantene på karpellen. Formen og størrelsen på stigmaet er forskjellig i forskjellige arter. Overflaten på stigmaet er svært ofte ujevn, tuberkulær og dekket med en klebrig væske, noe som bidrar til mer effektiv fiksering og oppfanging av pollen. I tillegg bærer stigmaoverflaten et tynt proteinlag, pellikkelen, som, i samspill med proteinene i pollenkornsporodermen, sikrer spiring av pollenrøret eller forhindrer det.
Kolonnen består av løst parenkymalt vev. Det ser ut til å løfte stigmaet opp, som er nødvendig for noen mekanismer i pollineringsprosessen. Morfologien til søylene er ganske mangfoldig og fungerer som et viktig systematisk trekk. Mange arkaiske familier (spesielt fra underklassen magnoliid) er preget av fraværet eller svak utvikling av stilen. Stiler er ofte uutviklet i mange spesialiserte vindpollinerte former, for eksempel i mange korn. I store vindbestøvede blomster (hos noen liljearter) når søylene en betydelig lengde, stigmaet bæres høyt opp, og dermed blir pollineringen lettere. Dette forlenger imidlertid banen til pollenrøret betydelig.
Eggstokken er den mest essensielle delen av pistillen, og bærer eggstokkene. Den er variert i form og utseende, som i stor grad bestemmes av typen gynoecium.
Festestedet til eggstokkene i eggstokken kalles morkake. Morkaken ser vanligvis ut som en liten hevelse, utvekst eller fremspring dannet av vevet i eggstokken.
Avhengig av funksjonene ved festingen av eggstokkene til eggstokkveggen, skilles flere typer placentasjon.
vegg, eller parental, når eggstokkene er plassert inne i eggstokken langs dens vegger eller på steder der fruktbladene er smeltet sammen.
aksial, eller aksial, når eggstokkene er plassert på den sentrale kolonnen av eggstokken, delt inn i reir i henhold til antall fruktblader.
fri sentral placentasjon, når eggstokkene utvikler seg på en fri sentral kolonne som ikke er forbundet med skillevegger til eggstokkveggen.
basal, når den eneste eggstokken er lokalisert helt i bunnen av en encellet eggstokk.
Typer gynecium:
1. apocarpous - fruktbladene vokser ikke sammen, og hver karpell danner en egen pistill (smørblomst, rose)
a) monomer - gynoecium består av 1 pistill, og den er dannet av 1 karpell (erter, plomme, kirsebær)
b) polymer - det er mange pistiller, men hver består av en fruktblad
(smørblomst, jordbær, villrose)
1. cenocarpous - pistillen er dannet av karpeller smeltet sammen
a) synkarpøs - karpeller vokser sammen med sideflater, flere ringer (tulipan) dannes. Det dannes flere reir inne i frukten.
b) paracarp - fruktbladene vokser sammen i kantene og danner en ring (valmue), eller det sentrale kammeret.
c) lysicarpous - karpellene vokser sammen i kantene, danner ett kammer eller hulrom, og en søyle stikker ut fra bunnen av eggstokken, som eggstokken er plassert på, og deretter frøene (nellik).
13. Eggløsning - en relativt kompleks formasjon, bestående av en frøstilk (kabelbane), som bærer en kjerne innelukket i ett eller to integumenter. Avhengig av arten utvikles fra én til mange eggløsninger på morkaken. Den utviklende eggløsningen består i utgangspunktet utelukkende av kjernen, men snart oppstår ett eller to integumentære lag (integumenter) med en liten åpning, en mikropyle, i den ene enden (fig. 6).

Ris. 6. Skjema for dannelsen av eggløsning og embryosekk.
1, 2, 3, 4 - utvikling av kjernen, isolasjon og meiose av archesporium-cellen, død av tre megasporer; 5, 6, 7, 8 - utvikling fra megasporen (resten) av den kvinnelige gametofytten - embryosekken.
På tidlig stadie Under utviklingen av eggløsningen vises en enkelt diploid megasporocytt i kjernen. Den deler seg mitotisk, og produserer fire haploide megasporer, vanligvis arrangert i en lineær tetrad. Dette fullfører megasporogenesen. Tre megasporer blir vanligvis ødelagt, og den fjerde, lengst fra mikropylen, utvikler seg til en kvinnelig gametofytt.
Den funksjonelle megasporen begynner snart å øke på grunn av kjernen, og kjernen deler seg mitotisk tre ganger. På slutten av den tredje mitosen er åtte datterkjerner plassert i fire grupper i to grupper - nær den mikropylare enden av megagametofytten, så vel som i den motsatte, chalazal, enden. En kjerne fra hver gruppe migrerer til midten av den åtte kjernecellene; de kalles polare. De tre kjernene som er igjen i mikropylarenden danner eggapparatet, bestående av et egg og to synergiske celler. I chalazal-enden dannes det også cellemembraner rundt kjernene som ligger her, og de såkalte antipodale cellene oppstår. De polare kjernene forblir i den binukleære sentralcellen. Denne åttekjernefysiske, syvcellede strukturen er den modne kvinnelige gametofytten kalt embryosekken.
En blomst er et komplekst organsystem som gir frøreproduksjon i blomstrende planter. Utseendet til en blomst i evolusjonsprosessen er en aromorfose, som førte til den brede distribusjonen av angiospermer, eller blomstrende planter, på jorden.
Blomsterfunksjoner:
- dannelsen av støvbærere med pollenkorn av karpeller (pistiller) med eggløsninger;
- pollinering;
- komplekse befruktningsprosesser;
- frø- og fruktdannelse.
Blomst- dette er et forkortet, modifisert og begrenset vekstskudd, bærende perianth, støvbærere, fruktblader (pistiller). Strukturen til blomster i alle blomstrende planter er lik, og formen er mangfoldig. Dette er manifestert idioadaptasjon - tilpasning til forskjellige måter pollinering.
Ytre struktur av en blomst
Blomsten ender hovedstammen eller lateral. Den bladløse delen av stilken under blomsten kalles pedicel. I fastsittende blomster er stilken fraværende eller sterkt forkortet. Pedikelen går inn i en forkortet akse av blomsten, dens stilkdel - beholder. Formen på beholderen kan være langstrakt, konveks, flat, konkav. Alle deler av blomsten er plassert på beholderen: begerblader og kronblader, støvbærere og pistil (pistil).
Begerbladene og kronbladene utgjør sammen perianth. Begerblader beskytter vanligvis blomsten, spesielt knoppen, mot skade, men de kan også tjene andre funksjoner. Fotosyntesen foregår i grønne begerblader som inneholder kloroplaster. Hos noen planter (tulipan, anemone) blir de kronbladlignende og fungerer som kronblader; kan tjene til å beskytte utviklende frukter, for deres distribusjon.
Begerblad stammer fra hest vegetative blader. Bevis på dette er deres morfologiske likhet med blader, tydelig uttrykt i noen planter (peon), og et spiralarrangement. Settet med begerblader danner et beger, som enten kan være separatbladet eller leddbladet.

kronblader funksjon for å tiltrekke seg pollinatorer og fremme vellykket pollinering. Opprinnelsen til kronbladene er todelt: hos noen planter er disse modifiserte støvbærere. Vannliljer har slike kronblader, så vel som representanter for familiene smørblomst, nellik, valmue, etc. En annen gruppe planter har kronblader, som begerblader, av bladopprinnelse (pion, magnoliaceae).
Samlingen av kronbladene til en blomst kalles Visp. Størrelsen, strukturen og fargen på kronen er variert, noe som er assosiert med pollineringsbiologien. Hos vindpollinerte planter er kronen enten underutviklet eller fraværende. Kronbladene kan vokse sammen i kantene og danne en høyhastighets krone (bindweed, petunia). I evolusjonsprosessen utviklet en slik krone seg fra en fritt kronblad.
Når en blomst har en beger og en krone, kalles perianth dobbel. Hvis det ikke er noen kronblad eller forskjellen mellom dem ikke er tydelig uttrykt, kalles perianth enkel. En enkel perianth kan enten være kroneformet med en lys farge - i tulipaner, liljer, liljekonvaller, eller koppformet, grønn - i hamp, quinoa, brennesler. Blomster uten perianth kalles nakne - i sir, selje.

Inne i perianth nærmere kronbladene er støvbærere. Antallet deres varierer fra én til ti eller mer. I utviklingsprosessen differensierte støvbæreren seg til filamentet og støvbæreren. Støvbæreren består av to halvdeler forbundet med en fortsettelse av filamentet. Hver halvdel av støvbæreren inneholder to sporangier, de kalles støvbærere, eller pollensekker.

Reirene er fylt med vev fra primære sporogene celler. Som et resultat av en serie påfølgende mitoser dannes mange moderceller, mikrosporer, fra primære sporogene celler. Modercellene deler seg deretter meiotisk, og danner tetrader av haploide mikrosporer. Hver slik mikrospore blir til et pollenkorn. For å gjøre dette øker den i størrelse og er dekket med et dobbelt skall: ytre (exine) og indre (intine). Det ytre skallet, på grunn av sin hovedkomponent - sporopollenin, er preget av høy motstand: det oppløses ikke i syrer og alkalier, tåler temperaturer opp til 300 ° C og forblir i geologiske forekomster i millioner av år.
Inne i pollenkornet dannes en mannlig gametofytt: den haploide mikrosporen deler seg mitotisk, og danner et større cellerør (vegetativt) og i det en liten generativ celle. Den generative cellen deler seg igjen mitotisk i to mannlige kjønnsceller - sædceller.

Den indre delen av blomsten er okkupert pistiller. Antallet deres varierer fra én til ti eller mer. Hver pistil er dannet av en eller flere sammenvoksede fruktblader.
På bunnen av pistillen - eggstokken - er eggene (ovules). Fra den øvre delen, i utviklingsprosessen, dannes en søyle som hever stigmaet over pistillen. I fravær av en stil kalles stigmaet sittende. Eggstokken kan være øvre hvis den er plassert på en flat eller konveks beholder, og alle andre deler av blomsten er festet under pistillen. I blomster med en nedre eggstokk vokser den konkave beholderen sammen med veggen, perianth og støvbærere er festet over pistillen.

Ved eggstokken til pistilen er det et hulrom - et rede. Det er en- og flercellede eggstokker. En flercellet eggstokk dannes som et resultat av sammensmeltningen av flere karpeller. Antall reir er lik antall sammenvoksede fruktblader. I hvert reir på eggstokkens vegger dannes eggstokker (ovules), enten fastsittende eller på frøstilker. Det er fra en (plomme, kirsebær) til flere tusen (valmue, orkideer).
Strukturen til eggløsningen (ovule)
I den anatomiske studien av eggløsningen skilles følgende komponenter ut:
- Funikkel;
- nucellus;
- dekker;
- mikropyle;
- embryosekk.
Av pedicel Næringsstoffer kommer inn i embryoposen og eggløsningen festes til eggstokken med den. Nucellus eggløsninger er et parenkymalt vev som nærer og beskytter megasporer. Utenfor er kjernen kledd i en eller to dekker(integumenter). De dekker ikke kjernen helt. Oftere, på toppen av eggløsningen, kobles de ikke sammen og danner et lite hull kalt mikropyle, eller polleninnføring.
Den innerste delen av eggløsningen er embryosekk, som hos angiospermer er den kvinnelige gametofytten.

Eggløsningen (ovulen) består av et makrosporangium og dets omkringliggende integument. I makrosporangium legges en modercelle, hvorfra en tetrad av haploide makrosporer dannes ved meiose. Tre av dem dør og blir ødelagt, og den fjerde (som gir opphav til den kvinnelige gametofytten) makrospore er sterkt forlenget i lengde, mens dens haploide kjerne deler seg mitotisk. Datterkjerner divergerer til forskjellige poler i den langstrakte cellen.
Videre deler hver av de dannede kjernene seg mitotisk to ganger til og danner fire haploide kjerner ved forskjellige poler i cellen. Dette er allerede en embryosekk med åtte haploide kjerner. Deretter, fra hver av de to firdoblene, sendes kjernene en etter en til midten av embryoposen, hvor de smelter sammen og danner en sekundær diploid kjerne.
Etter det oppstår cellepartisjoner mellom kjernene i cytoplasmaet til embryoposen og den blir syvcellet.
Ved en av polene til embryoposen er eggapparatet, som består av et større egg og to hjelpeceller. På motsatt pol er det tre antipodale celler. Alle seks cellene er haploide. I midten er en diploid celle med en sekundær kjerne.
Hos de fleste planter har blomstene støvbærere og pistiller og kalles bifile. Det er blomster og samme kjønn: staminate (mann) eller pistillat (kvinne). Mannlige og kvinnelige blomster kan lokaliseres på samme individ, en slik plante kalles monoecious (agurk, mais, eik, bjørk), og hvis på forskjellige individer - toebolig (hamp, pil, poppel). Enkjønnede blomster og toboplanter er en av tilpasningene for krysspollinering.
Diagrammer og fora over planter
Til Kort beskrivelse blomster bruker diagrammer og formler. Et diagram er en skjematisk projeksjon av elementene i en blomst på et plan vinkelrett på dens akse. Alle deler av blomsten, bracten og morsskuddet er indikert med visse ikoner: begerblader - med en krøllete parentes, kronblader - med en parentes, støvbærere - med et tverrsnitt gjennom støvbæreren, pistill - med et tverrsnitt gjennom eggstokken .
Ved kompilering av blomsterformelen er perianth betegnet med bokstaven O, begerblader - H, kronblader - L, støvbærere - T, pistill - P. Antallet blomsterdeler er angitt med et tall skrevet i bunnen av bokstaven. Hvis det er mer enn 12 støvbærere og pistiller, sett et ikon - ∞. Når deler av blomsten vokser sammen, er de tilsvarende tallene tatt i parentes. Den øvre eggstokken er indikert med en horisontal linje under nummeret, den nedre - over antall pistiller.
Bifile og enkjønnede blomster -Bifile og enkjønnede blomster
Blomster er biseksuelle (med androecium og gynoecium) eller enkjønnede (bare med androecium eller bare med gynoecium). Blomster av samme kjønn kan enten være på samme plante, som i eik, bjørk, melkegress, mais (og så er planten som helhet bifil), eller på forskjellige planter, som i poppel, selje, hamp (da har vi hanner) og hunnplanter). I denne forbindelse har to begreper lenge eksistert i den botaniske litteraturen - monoecious og dioecious. Siden Linnés tid har mange botanikere brukt disse begrepene på plantene selv og snakker om tobolige og eneboende planter. Hvis både bifile og enkjønnede blomster finnes på planten, som i mange Compositae, så sies de å være polygame (fra det greske poly - mange og gamos - ekteskap). Fra og med O.P. de Candoll, S.L. Zndlihor, D. Weptam og J.D. Hooker, og slutter med A. Engler, R. Wettgatein, A.B. Repdl og J. Hutchinson, bruker mange forfattere begrepene "dobbelt", dvs. "enebolig" kun for blomster, ikke hele planter. Tvistene som noen ganger oppstår om hvilken av disse to brukene av begrepene som er mer korrekt, er i hovedsak meningsløse. Cannabis eller selje kan like gjerne sies å være toeboe eller å ha toebolige blomster. Avhengig av konteksten kan den ene eller den andre bruken av disse vilkårene være mer praktisk, og dette vil ikke i noe tilfelle føre til misforståelser.
Det er all grunn til å tro at enkjønnede blomster oppsto fra bifile, og i enkjønnede blomster er dioecy tydeligvis senere enn enkjønnet. Siden andre halvdel av forrige århundre har en rekke studier på den komparative morfologien og biologien av pollinering ført til konklusjonen at enkjønnede blomster oppsto fra tokjønnede blomster som et resultat av underutvikling eller fullstendig undertrykkelse av støvbærere i noen tilfeller, og bærblader i andre. I likekjønnede blomster av mange slekter og hele familier er det ofte bevart reduserte rester (rudimenter) av støvbærere og fruktblader (de såkalte staminoder og karpeller). Slike restformasjoner kan sees i blomstene til representanter for forskjellige familier, inkludert platantreet, noen morbær, brennesler og valnøtter. Grunnleggende biologisk årsak overgangen av bifile blomster til enkjønnede er mer pålitelig krysspollinering, som påpekt av Charles Darwin.
Etter å ha lest disse linjene, kan leseren stille spørsmålet: er det mulig å snakke om felten til en blomst, fordi en blomst er en del av en sporofytt, eller aseksuell generasjon, og dermed er blottet for sex? Noen botanikere mener det, og i stedet for begrepene «mann», «kvinnelig» og «bifil» foretrekker de å bruke begrepene «utholdelig», «pistillat» og «perfekt» (perfekt i den forstand at det finnes både støvbærere og fruktblader). Imidlertid fortsetter de fleste botanikere å bruke begrepene bifil og samme kjønn, mann og kvinne, og med god grunn. Morfologisk er blomsten utvilsomt en del av sporofytten, men funksjonelt er den mest direkte relatert til den seksuelle prosessen.
Når vi snakker om mannlige og kvinnelige blomster, mener vi deres rolle i forberedelsen av seksuell reproduksjon, og ikke tilhører den seksuelle generasjonen (gametofytt). Saken er at den genetiske og fysiologiske differensieringen mellom det mannlige og kvinnelige kjønn også går over til den aseksuelle generasjonen, en viss seksualisering av sporofytten skjer. Dette er spesielt uttalt hos tobolige planter (planter med tobolige blomster). Hann- og kvinnelig cannabisplanter er genetisk og fysiologisk forskjellige, og man kan til og med si at mannlig cannabis ikke er mindre maskulin enn hanndyr. Av samme grunn kan vi betrakte støvbæreren som en hannstruktur, og fruktbladen som en hunn.
Planteliv: i 6 bind. — M.: Opplysning. Under redaksjon av A. L. Takhtadzhyan, sjefredaktør corr. USSR Academy of Sciences, prof. A.A. Fedorov. 1974 .
Magnoliidae-underklasse (Magnoliidae) Magnoliidae-underklasse (Magnoliidae) De mest arkaiske levende blomstrende plantene introduseres i underklassen Magnoliidae. Hoved- og sentralgruppen av magnoliider, magnolianorden, utmerker seg ved en spesiell primitivitet. Men primitiviteten til individuelle
Hamamelis-familien (Hamamelidaceae) Hamamelis-familien (Hamamelidaceae) Vårt bekjentskap med Hamamelis-ordenen, vil vi begynne med Hamamelis-familien, som inntar en nøkkelposisjon i den. Dette er en eldgammel familie, den tiden for den største velstanden var tertiærperioden. som show
Klassifisering og fylogeni av blomstrende planter Klassifisering og fylogeni av blomstrende planter De første forsøkene på å klassifisere blomstrende planter, samt flora generelt var basert på noen få, vilkårlig tatt, lett iøynefallende ytre tegn. Dette var rent kunstige klasserom.
UNDERKLASSE DILLENIIDAE (DILLENIIDAE) UNDERKLASSE DILLENIIDAE (DILLENIIDAE) Dilleniidae er en av de største underklassene av blomstrende planter. I fylogenetiske termer er det også en av de viktigste grenene av slektstreet, og er en kobling mellom magnoliider og rosider. På
UNDERKLASSE ROSIDS (ROSIDAE) UNDERKLASSE ROSIDS (ROSIDAE) Bestillingene som inngår i underklassen Rosids er svært forskjellige i utseende, strukturen til blomsten og anatomien til de vegetative organene. Imidlertid er de forent av en felles opprinnelse og, som resten av underklassene, representerer rosider en naturlig
LILIID UNDERKLASSE (LILIIDAE) LILIID UNDERKLASSE (LILIIDAE) Liliider er en stor underklasse av monocotyledoner, inkludert alle de største familiene (unntatt palmer og aronnikovye, som tilhører den neste underklassen av arecida). Blant liliidene er det begge relativt primitive planter, sammenlignbare i
POLYGAM POLYGAM - et fenomen der bifile og enkjønnede blomster finnes på samme plante eller på forskjellige eksemplarer av samme art. Ordbok over botaniske termer. - Kiev: Naukova Dumka. Under hovedredaksjon av d.b.s. I.A. Dudki. 1984.





